Fiordland penguin

Fiordland penguin / tawaki
Eudyptes pachyrhynchus
| Thomas Mattern & Kerry-Jayne Wilson | Document date: 12 April 2019; DOI: 10.36617/SoP.tawaki.2019-04 |
Summary
The Fiordland penguin or tawaki (Eudyptes pachyrhynchus) is the only crested penguin species to breed on the New Zealand mainland and, therefore, the only crested species in the world that lives in relative proximity to human settlements. Although only breeding in remote parts of the mainland and satellite islands in South Westland, Fiordland and Stewart Island that are mostly difficult of access tawaki are exposed to many anthropogenic influences including inshore fisheries, unregulated tourism, and pollution, that other crested penguins are not. The species is believed to have undergone a significant decline in recent decades, although it is unclear whether this trend continues. Currently, the official population estimates for the species range between 5,500 and 7,000 mature individuals1, although recent surveys suggest that tawaki are more common than generally thought. Until recently little was known about the species marine ecology; diet studies carried out in the 1980s found that the penguins fed predominantly on cephalopods and krill in an open coast environment, while fish dominated the diet in the Foveaux Strait region. Studies conducted since 2014 have significantly expanded our knowledge about the species.
Previous reviews of biology and priority lists
The first detailed account of tawaki was published by John Warham2 which primarily focussed on terrestrial aspects of species biology. A comprehensive, encyclopaedic review was compiled by Marchant & Higgins3 which cites several sources that cannot be accessed today. A report summarised the results of the Department of Conservation’s monitoring programme and identified several issues related to survey design4. More recently a review of available knowledge largely based on unpublished reports was compiled by Mattern5; that chapter serves as basis for this review.
Research and conservation priorities for tawaki have been listed in Taylor6 and Mattern5. Both stress the urgent need for a better understanding of the species’ demography and marine ecology in order to improve conservation outcomes.
Here, we focus on aspects of the species’ biology that are deemed most important for conservation. Below we list topics and relevant publications that will not be discussed in detail in this review.
- Genetic & Social monogamy7
- Vocalisation8
- Egg formation9 & reproductive endocrinology10
- Brood reduction11,12
Conservation status
The Department of Conservation lists tawaki as ‘Nationally Vulnerable’ (criteria D, 5,000-20,000 mature individuals, predicted decline 10-50%13). The IUCN red list classified the species as ‘Vulnerable’ due to the species’ small population that underwent ‘rapid decline over the last three generations’1.
Taxonomy
Fiordland penguins have only recently been recognised as distinct from the Snares penguins (Eudyptes robustus). In the 1970s, the Checklist of New Zealand Birds considered Snares, Fiordland and even Erect-crested (E. sclateri) penguins as conspecifics14. This was disputed due to morphological and ecological differences between each taxa15–17. More recently, studies that examined relationships using morphology, protein data, and mitochondrial and nuclear DNA have all supported the recognition of three separate species18–21. Nevertheless, a review of the taxonomic status in 2008 still considered Fiordland and Snares penguins as conspecific22. Consultations by Birdlife International in 2014 led to the calculation of the Tobias score23, which demonstrated that tawaki and Snares penguins should be considered separate species (see http://bit.ly/tawaki-tobias-score).
Distribution
The breeding range of tawaki is limited to the south-western coast of New Zealand’s South Island where the penguins occur in lose colonies in coastal rainforests, sea caves or under rock boulders of rocky shores. Mattern5 provides an extensive list of tawaki breeding sites that have been surveyed in the past. Penguin numbers at 43 colonies have been estimated sometime between 1991 and 2009. On the mainland, tawaki range from Heretaniwha Point near Bruce Bay, South Westland (S43.59, E169.55) to Coal Island in southern Fiordland (S46.12, E166.63). In Foveaux Strait, the penguins breed on Solander Island/Hautere, Codfish Island/Whenua Hou, Stewart Island/Rakiura and its outliers5.

Figure 1. Tawaki in the Harricon Cove, Milford Sound/Piopiotahi breeding colony which features a typical rainforest setting. (Photo: Thomas Mattern)
Tawaki have been recovered from a number of natural fossil deposits and archaeological middens throughout coastal New Zealand, and there are numerous publications that summarise findings24,25. Although difficult to distinguish from other crested penguins (and even Megadyptes penguins), most prehistoric specimens found to date on the New Zealand mainland were tawaki21. The large number of bones excavated from many sites in coastal New Zealand indicate that tawaki have probably inhabited New Zealand at least since sea level stabilised, 6000 years ago.
It has been suggested that historically tawaki had a wider distribution with breeding sites in the north of the South Island and potentially even the lower North Island5. However, recent studies seem to contradict this idea. A study looking at genetic information in prehistoric bones, museum skins and blood samples found that the genetic diversity of tawaki does not suggest any substantial range restriction over the last thousand years21. Moreover, while historic samples were obtained from adult penguins found as far north as the North Cape, no juvenile bones or eggshells were excavated from the upper South Island or lower North Island. This may indicate that bones from these northern sites originate from vagrants rather than resident breeders. This hypothesis is further supported by a recent examination of the pre-moult dispersal in tawaki26. The authors argue that genetic predisposition may drive the penguins to travel to the sub-Antarctic front some 2,000 km south of New Zealand and that the current range of the species likely represents the geographic extremes at which this strategy is viable.
Outside the breeding season, recent satellite tracking data suggests that tawaki spend most of the time prior to their moult in sub-Antarctic waters between 1,500 and 2,500 km south-east of the South Island26. Outside the breeding season, tawaki (mostly moulting birds) have been observed around most of the South Island, but also on the Snares/Tini Heke, Auckland, Campbell and Macquarie Islands. They are common visitors to Tasmanian shores and are occasionally reported from southern West Australia all the way to New South Wales; there is one unconfirmed report of a tawaki on the Falkland Islands3. Where tawaki go after completing the moult is currently being investigated; it appears that birds tend to show similar dispersal movements to the pre-moult period (Mattern, unpublished data).
Numbers and population trends
A series of distribution surveys were carried out between 1990 and 1995 that covered most of the species’ current breeding distribution27-31. To date, these surveys represent the only attempt at an estimate of tawaki numbers. The authors concluded that there were 2,500-3,000 breeding pairs31. However, methodology and scope of these surveys made undercounting of penguins highly likely5. Using accounts published prior to 2016, a review of Fiordland penguins for the IUCN Red List concluded that there are between 2,500 and 9,999 mature individuals1.
More recently, nest counts carried out in Fiordland32 and South Westland 33 found considerably more penguins than reported by the survey 20 years earlier. A ground survey conducted in Milford Sound/Piopiotahi in the spring of 2016 found 77 nests and resulted in an estimate of between 130 and 150 breeding pairs in this fjord32, a stark contrast to the 9 nests found in the 1990s27. Similarly, nest searches carried out in the spring of 2014 between Cascade River and Martins Bay found a total of 835 nests33; the previous estimate for that region was just 150 nests31.
The estimates from the 1990s and recent surveys either indicate significant undercounts in the earlier surveys or suggest an increase in tawaki numbers in the past 20 years. McLean et al.31 themselves caution readers to consider their counts to be “a minimum estimate” due to the logistic constraints of their survey. So, undercounting certainly contributes to the apparent differences in penguin numbers between the 1990s and the recent surveys.
Most recent estimates of tawaki numbers32,33 can be used to adjust counts from the 1990s surveys. Along the southern West Coast Robin Long33 counted 850 breeding pairs while McLean et al.31 only recorded 150 pairs in the early 1990s suggesting a population increase (or previous underestimation) of 566%. Similarly, the recent nest counts provided for the Milford Sound population of 77 breeding pairs32 compared to 9 breeding pairs recorded there in 1990s5 would represent 770% increase in numbers. Assuming that much higher penguin numbers apply to the species’ entire breeding range and applying a conservative adjustment factor of 500% to the IUCN numbers, the current population size of tawaki could range between 12,500 and 50,000 mature individuals.
Yet, a recent paper analysing tawaki population trends using data from DOC’s monitoring programme that operated from 1990 to 2010 concludes that the tawaki population is still in decline34, in line with the assessments in Taylor6 and the IUCN red list1.
However, other observations seem to suggest numbers have actually increased:
- Beach counts (n=1,270 counts) of tawaki commuting between the sea and their breeding colonies at a beach north of Haast conducted over the past 20 years showed an increase in penguin numbers from an average 8 birds per observation in 1996 to 12 birds in 2017 (Gerry McSweeney, unpubl. data)
- Other credible observers have noted an increase in tawaki numbers in recent decades. Several tourist boat skippers operating in Milford Sound during the past two decades believe that penguin numbers have increased; as do some fishermen operating in Dusky Sound. Tawaki were seldom seen along the north-east coast of Stewart Island in the 1970s (K.-J. Wilson, pers. obs.), where in 2005 and 2014, they seemed to be omnipresent, both at sea as well as on land (Mattern, pers. obs., see https://www.tawaki-project.org/2014/11/07/little-cave-men/).
- Recently tawaki have attempted to breed in the Catlins which may suggest a range expansion for the species35.
- Genetic analyses of 72 historic and recent tawaki samples found no evidence for significant changes in population size for tawaki over last thousand years21.
The assumption by Taylor6 that numbers were in decline was based on observations from a single site, Open Bay Islands. Between 1988 and 1995 the tawaki population on the island reportedly declined by 33%36. However, during that period research that involved handling of eggs shortly after laying and daily nest checks was undertaken on the island 11. As tawaki are perhaps the most timid of all crested penguin species37, that intensive research may have contributed to this apparent decline.
In this light it seems doubtful that negative population trends observed at Open Bay Islands are representative across the species’ entire range. The information available does not allow us to determine population trends with any degree of certainty.
The diverse breeding and foraging habitats occupied by tawaki are reflected in notable differences in their ecology and demographic parameters. For example, while the breeding success and number of breeding tawaki from Jackson Head, West Coast was significantly impacted first by a strong El Niño in 2015 and then by an invasion of stoats (Mustela erminea) in 2016, the situation in Milford Sound was diametrically different with above average breeding success and stable nest numbers38,39. Similarly, breeding success was found to be high and penguin numbers stable at Codfish Island/Whenua Hou in both 2016 and 201739. At some sites, terrestrial predators can have serious impacts on breeding success while at other sites these predators are absent39. Predation by stoats varies season to season and site to site. At Jackson Head no predation events were recorded during the 2014, 2015 and 2017 seasons, yet all breeding attempts failed in 2016 with stoat predation being the likely main cause; at the same time no predation events were recorded at their Gorge River study site 45 km away40.

Figure 2. Impenetrable tawaki habitat in a kiekie patch on Breaksea Island. Yellow arrow indicates tawaki nest. (Photo: Thomas Mattern)
Tawaki are notoriously difficult to monitor. They mostly breed in inaccessible places such as in thick kiekie (Freycinetia banksia) vegetation, narrow rock crevices or labyrinthine caves making it very difficult to conduct reliable counts 5,32 (see also https://youtu.be/Z2Qc6SrGDjc). Observer experience and endurance is of utmost importance to achieve reproducible monitoring results and can significantly affect the reliability of ground counts. For example, on three consecutive surveys an increasing number of nests were found in previously overlooked areas even though the observers were highly experienced and knew the terrain extremely well33,41,42.
Overall, it is unlikely valid conclusions about population trends can be drawn from ground counts. The penguins’ cryptic breeding habits and often impenetrable breeding habitat effectively prevents the application of traditional nest searches to obtain reliable information on population trends. Instead, a focus on determining key demographic parameters (i.e. survival rates, fecundity) from mark-recapture studies across their varied habitat should be used to assess population trajectories.
Demography
Between 1990 and 2010 the Department of Conservation conducted annual nest counts at various sites ranging from Codfish Island/Whenua Hou, Foveaux Strait to Monro Beach, South Westland34. More recently, double counts (the count repeated by a second team two days later) at selected sites were employed to minimize the potentially substantial observer error from the single counts prior to 2010; this indeed delivered more reliable population estimates37. However, limited resources have since led to a cessation of the DOC monitoring programme.
Based on the monitoring data, Otley et al.43 determined core demographic variables for the species. Survival probabilities were calculated to be around 89% for adult penguins, with juvenile survival – defined as survival until first breeding – estimated at 77%. Combined with a mean breeding success of 0.61±0.02 chicks per pair, tawaki core demographic variables appear to be higher than for most other crested penguin species44–46.
The Tawaki Project has researched the marine ecology of tawaki at three sites – Jackson Head, Milford Sound and Codfish Island/Whenua Hou – and in conjunction with this work has monitored breeding success. While breeding success at Jackson Head varied due to the aforementioned impacts of El Niño and stoat invasion between 0.12 and 0.94 chicks per pair (2014-2017), reproductive efforts of tawaki from Harrison Cove, Milford Sound were consistently high (0.8 - 1.1 chicks per pair, 2015-2017); breeding success was similarly high at least until crèching on Codfish Island/Whenua Hou (Mattern et al. unpubl. data). However, as with ground counts, determination of breeding success is difficult as chick survival can only be determined with certainty until the crèching stage during which chicks are highly mobile and often impossible to locate.
Breeding biology
Except for a few open nests, tawaki are cryptic breeders that prefer to nest in inaccessible locations. Along open coast sites like Jackson Head, the penguins tend to breed in dense, impenetrable kiekie shrub often in small clusters of nests2. At Gorge River, the penguins breed in a more open forest setting, primarily under tree roots or fallen trees, in amongst kiekie or tangles of supplejack (Ripogonum scandens)33. In Harrison Cove, Milford Sound less than a third of all nests are under rock overhangs or upturned tree roots. Most of the nests are in rock crevices or in cavities and caves under rock falls. At Sinbad Gully, Milford Sound, the penguins occupy a steep slope that is dominated by windfall of rimu (Dacrydium cupressinum) and other large trees; a few kilometres up the fjord, the majority of the penguins nest in an extensive warren under remnants of a glacial moraine32. On Codfish Island/Whenua Hou, the penguins breed in dense tree fern groves, mainly in dug out, deep burrows in the soft peaty soil or in deep hollows under tree roots, while along the north-east coast of Stewart Island the penguins breed in sea caves and fissures in cliffs only accessible from the sea (Mattern, pers. obs.).
Nests usually consist of shallow bowls lined with twigs and stones (Fig. 3).

Figure 3. Tawaki nest in Harrison Cove, Milford Sound/Piopiotahi (Photo: Thomas Mattern)
John Warham2 published the first and seminal account of tawaki breeding biology, although his observations were largely limited to a single site, Jackson Head. In the late 1980s and early 1990s, St. Clair11,47 focussed on site fidelity and incubation behaviour in an effort to decipher the crested penguins’ obligate brood reduction. Some aspects of the breeding biology of tawaki are also touched on by Ellenberg et al.37.
Mattern5 provides a detailed summary of the species breeding biology thus only a condensed account is presented here.
Tawaki return from three months at sea to their breeding colonies in mid-June, with the majority of penguins returning to the nest sites they used in the previous year; about two-thirds of the penguins reunite with their previous partner2,47. Egg laying occurs over a 10-day period in late July and early August, the clutch size is two with the eggs being laid 3-6 days apart. As with other crested penguins, the first-laid A egg is smaller than the second-laid B egg, although the size difference is less marked than in other crested penguins48. While largely synchronized within colonies, the timing of egg laying seems to vary between sites by as much as 2-3 weeks37.
In tawaki, the roles during incubation are the reverse of that in other crested penguins. The pair spends the first 5-10 days after the B-egg is laid together at the nest, the female then leaves on a two week long foraging trip2; in all other crested penguins it is the male that makes the first foraging trip48. After the females return, the male penguins leave for up to two weeks usually returning a few days prior to egg hatching. Like all other crested penguins, tawaki are considered obligate brood reducers that generally only raise one chick. However, McLean et al.7 noted that in years when food is abundant, up to 12% of tawaki pairs can fledge both chicks. Some tawaki breeding in Milford Sound/Piopiotahi and on Codfish Island/Whenua Hou can also fledge both chicks38,39,49.
Eggs hatch throughout September, 31-36 days after the B-egg is laid2,11. In nests where only one chick is raised, the smaller chick that hatched from the A-egg usually dies within a week after hatching, severely disadvantaged by smaller body size compared to its sibling. Both Marchant & Higgins3 and St Clair50 state that 50-60% of nests only hatch one egg; the rate of egg loss was lower during recent studies38,39,49,50. The male penguins guard the chicks for up to three weeks, while the female is the sole provider of food to her offspring2.
From early October, chicks may form small crèches that are often guarded by one male penguin. Mostly, however, both male and female undertake foraging trips at this stage. It is mainly the female that feeds the chicks, although males increase their food contribution closer to fledging2. Chicks fledge between mid-November and early December. The parents leave on their pre-moult foraging trips shortly after the chicks have fledged.
Moult
Satellite tracking of adult tawaki from Gorge River, South Westland during the pre-moult dispersal November 2016 to February 2017 found that the penguins travelled extraordinary distances compared to other crested penguins at this stage of the annual cycle26. The birds left their colonies between mid-November and early December with those birds departing earlier travelling towards the subtropical front some 800-1000 km south of Tasmania, while penguins leaving in December ventured further south to the sub-Antarctic front southeast of Macquarie Island. Return journeys ranged from 3,500-6,800 km during the 8-10-week sojourn at sea before returning to the mainland to moult. Of the five birds that could be tracked for the entire trip, three penguins returned to their breeding colony at Gorge River to moult. The remaining two birds moulted at Dusky Sound and Sutherland Sound, respectively. Warham (1974) states that penguins generally return in mid-January and early February to moult. The satellite tracking study found that some birds may return as late as the end of February26.
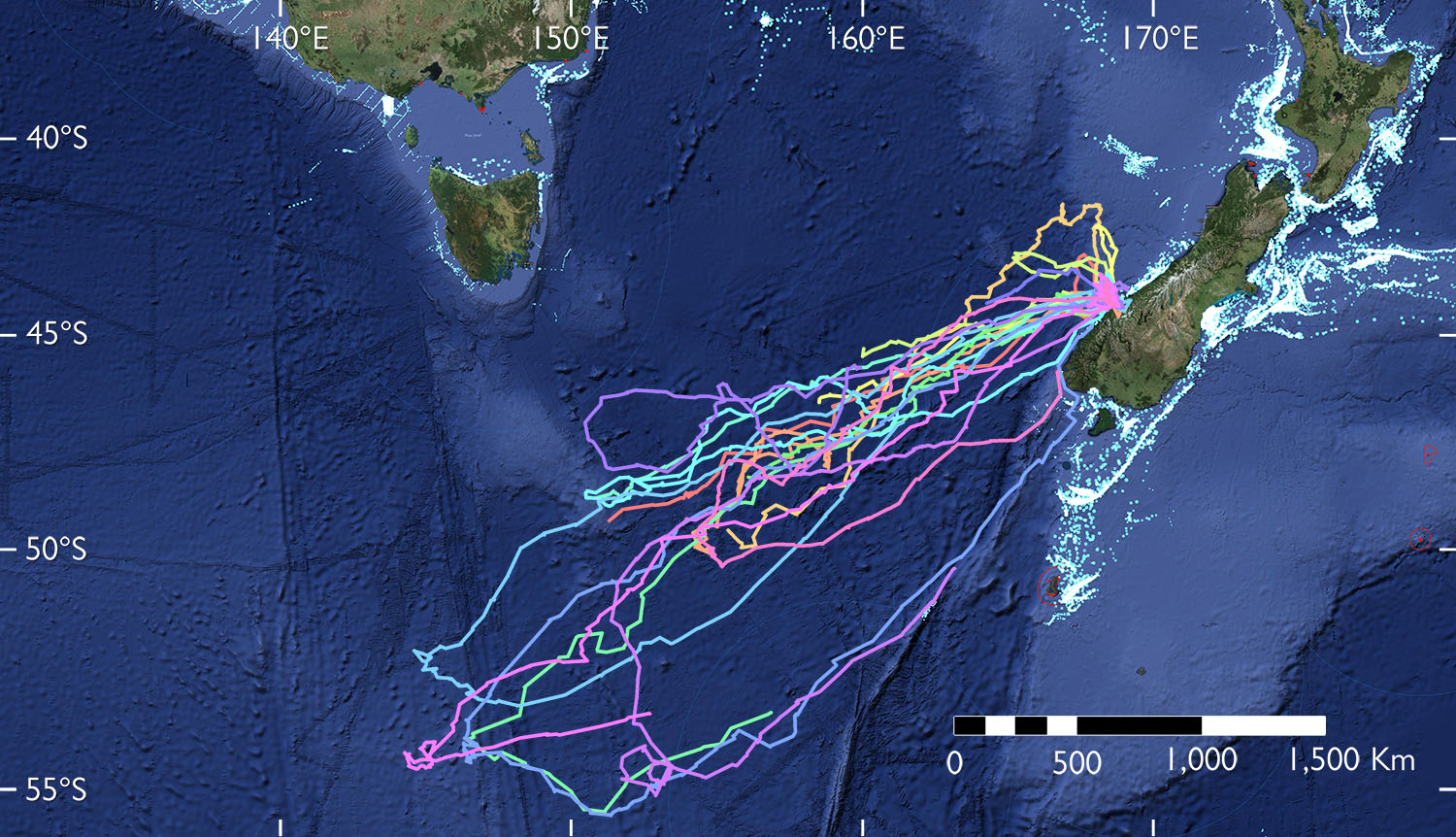
Figure 4. Pre-moult dispersal of tawaki from Gorge River, Southwestland December 2016-February 2019 as determined by satellite tracking; graph adapated from Mattern et al. 26
Although it has been stated that tawaki generally return to their breeding colonies to moult2, this seems to vary between sites. For example, tawaki colonies on the western side of Jackson Head were found to be virtually devoid of moulting penguins or sign that penguins had recently completed moulted there in February and March 2014-2018 (Mattern, pers. obs.). Instead, moulting tawaki are commonly seen in the nearby settlement of Neils Beach and along the coastal road to Jackson Bay (Geoff Robson, Greenstone Helicopters, pers. comm.). In Harrison Cove, Milford Sound/Piopiotahi (Fig. 5), on the other hand, there seem to be more penguins moulting there than are present during the breeding period. On Codfish Island/Whenua Hou, the colonies seem to be largely occupied by moulters that had bred there months earlier (Mattern, pers. obs.).
Replacement of feathers takes roughly three weeks; the penguins then depart on their winter sojourns2.

Figure 5. Tawaki undergoing the moult in Harrison Cove, Milford Sound/Piopiotahi (Photo: Thomas Mattern)
Food and foraging
Until recently the only information available on the marine ecology of tawaki was derived from two studies of their diet in the early 1980s52,53. Prey taken varied between regions with arrow squid (Nototodarus sloani) (relative biomass contribution: 85%) and krill (13%) dominating the diet of tawaki from Martins Bay, Fiordland53, while fish (85%), especially larval stages, were most important for penguins from Codfish Island/Whenua Hou, where cephalopods and crustaceans were minor constituents of their diet52. More recently, the prey composition of penguins from Jackson Head was investigated by identifying prey DNA in penguin scats (Julie McInnes et al., unpubl. data). Fish were the most common prey category (frequency of occurrence: 75%) followed by cephalopods (15%) and krill (10%). Interestingly, jellyfish DNA was found in several of the samples which could indicate that, like yellow-eyed penguins54, tawaki may feed on fish larvae associated with jellyfish.
Since the 2014 breeding season, The Tawaki Project has studied the foraging behaviour of chick rearing tawaki at sites representative of the species’ varied marine habitat, i.e. Jackson Head, South Westland (continental shelf); Harrison Cove, Milford Sound/Piopiotahi (fjord); and Codfish Island/Whenua Hou, Foveaux Strait (shallow, coastal). The at-sea movements and diving behaviour differed significantly between these sites.
At Jackson Head, the penguins generally foraged between 10 and 50 km offshore, mostly north-west of the colony. Foraging ranges and trip durations were influenced by prevailing environmental conditions travelling further during longer duration trips during years dominated by southerly winds (e.g. El Niño conditions) while north-westerly winds appear to create more favourable foraging conditions closer to the coast (https://youtu.be/1c4B9sdL8Y0?t=12m50s).
During three years of research in Milford Sound/Piopiotahi, there has been just one observation of a penguin leaving the fjord. All other data clearly show that while feeding chicks tawaki forage exclusively within the fjord; most of the foraging activity occurring just 1-4 km from their breeding colony. It is noteworthy that the penguins tend to spend most of their time along the northern side of the fjord within one of New Zealand’s few marine reserves (Fig. 6).
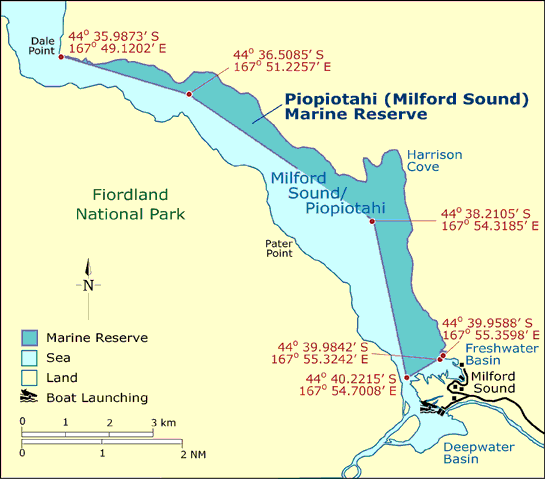
Figure 6. Tawaki undergoing the moult in Harrison Cove, Milford Sound/Piopiotahi (Source: http://bit.ly/milford-sound-MPA)
Tawaki from Codfish Island/Whenua Hou tracked in the 2016 and 2017 breeding seasons and have shown two types of foraging strategies. Birds either forage very close inshore following the coast line, presumably targeting prey in the kelp forests, or they travelled 20-30 km to the west where the shallow Foveaux Strait seafloor drops away into the deep-water Solander Trough (Mattern & Ellenberg 2017; Mattern et al. unpubl. data).
Overall, it appears that penguins from South Westland show the greatest foraging effort and are most susceptible to environmental perturbations. What this means for tawaki breeding along the open Fiordland coast – especially populations in interface regions where birds can choose to forage inside or outside the fjords (e.g. Breaksea Island, Shelter Islands) needs to be addressed.
Diving behaviour of tawaki is also being studied as part of The Tawaki Project. A maximum dive depth of 99 m has been recorded for a female from Jackson Head. However, most dives are considerably shallower, seldom exceeding 30 m, indicating that tawaki mostly feed close to the surface. Body acceleration has also been recorded which allows a more detailed analysis of the penguins’ diving behaviour55. Detailed analysis of all diving data is pending.
Predators
Although Warham2 states that seals are unlikely to play an important role in predation of tawaki, recent observations show that tawaki are taken by fur seals (Arctocephalus forsteri) in Milford Sound (Daniel Crook, Southern Discoveries, pers. comm.). Moreover, groups of fur seals apparently undertaking coordinated pursuit of tawaki have been observed at the entrance of the fjord (Turgut Ortabas, Southern Discoveries, pers. comm.). Dead tawaki have been found on beaches with obvious signs of shark bites (Fig. 7, Mattern, pers. obs.). However, beyond this little is known about predation of tawaki at sea.

Figure 7. Dead tawaki found on a Southwestland beach 2014 with rows of laceration from shark bites running along one flipper and its torso (Photo: Thomas Mattern)
On land, stoats have a significant impact on breeding success during some years at some locations. At Jackson Head and Gorge River they have been observed to take eggs and kill tawaki chicks39,40. In the 2016 breeding season, an invasion of stoats apparently caused almost complete breeding failure on the western side of Jackson Head. Yet in 2014, 2015 and 2017 no predation events were recorded at that colony40. Conversely at Gorge River, 45 km away, where nests were also monitored by motion activated cameras, one egg was taken by a stoat in both 2014 and 2015 but no predation events were recorded in 2016, the year stoats caused breeding failure at Jackson Head40. The 2016 breeding season followed a mast event in the podocarp forests near Jackson Head and the stoat invasion may have been a result of stoats moving onto the headland as food availability in the neighbouring forests declined. Disappearance of chicks at Jackson Head in 2016 only ceased after traps had been deployed and three stoats killed. A network of self-setting traps has since been established on Jackson Head. The effectiveness of ongoing predator control by the Wilderness Lodge Moeraki and the Department of Conservation at one site north of Haast using brodifacoum bait stations (until 2006) followed by regular 1080 drops is likely to be linked to increasing penguin numbers determined from beach counts during the past 20 years (Gerry McSweeney, pers. comm.).
The endemic weka (Gallirallus australis) preys upon tawaki eggs and chicks6,12,50. Weka have been introduced to some islands where tawaki breed including Open Bay Islands and Solander Island6. However, Mattern5 suggests that if the impact of weka was severe, tawaki populations at those sites would probably have declined in the immediate years following weka introduction; yet both tawaki and weka remain common on both.
Taylor6 suggested rats (Rattus sp.), and brushtail possums (Trichosurus vulpecula) to be potential predators of tawaki eggs and chicks. Rats, mice and brushtail possums are present in the Jackson Head and Gorge River tawaki colonies, they frequently pass by and occasionally investigate tawaki nests but none have been seen preying on tawaki eggs, chicks or harassing adult penguins40. Rats and possums may scavenge expelled eggs and dead chicks (Mattern, pers. obs.). Dog attacks have been reported at some accessible breeding sites including Jackson Head (Jacinda Amey, DOC Haast, pers. comm.).
Human disturbance through research and unregulated tourism is believed to have a significant impact on tawaki6. While people’s presence at landing sites do indeed cause delays when penguins want to come ashore, the penguins seem to be more tolerant to research interactions than previously expected51. This is presumably due to the penguins’ attachment and investment in their nest rather than a greater tolerance of disturbance37. Unregulated visits to the colony at Munro Beach, South Westland has apparently led to a decline in the numbers of tawaki breeding there although robust data to support this is lacking. Along the Jackson Bay Road, tawaki occasionally get run over by vehicles, particularly during the moult (Geoff Robson, Greenstone Helicopters, pers. comm.)
Threats
Comprehensive summaries of threats have been published in Mattern 5 and Otley et al. 34, however neither provides a threat level ranking or considers regional differences. In this report, we rank threats into major, medium and minor categories and provide a regional context.
Environmental disasters
Earthquakes
South Westland: Major
Fiordland: Medium
Foveaux Strait: Medium
Mattern5 notes the high level of geological activity throughout the tawaki breeding range. Geological modelling found that a major earth quake (magnitude 8 or higher) in South Westland is likely to occur in the coming decades and this will have severe effects coastal habitats (Biasi et al.56, see also http://projectaf8.co.nz/). When such an earthquake occurs, it will affect most if not all of the tawaki breeding range, although the nesting habits of tawaki on Stewart Island and Codfish island/Whenua Hou where shaking will be less severe makes them less vulnerable to violent earth movements.
A large-scale earthquake would have the greatest impact if it were to strike during the breeding season, particularly nest formation and early incubation (June-July) and during the moult (February) when most adult penguins are in their colonies.
Pollution
Oil spills
South Westland: Medium
Fiordland: Major
Foveaux Strait: Major
Oil spills caused by shipping disasters would likely have a localized impact in the vicinity of the wreckage. The oil spill caused by the wreck of the container vessel Rena severely affected some 20-30 km of coast in the Bay of Plenty 57, although the impact could have been more widespread if more oil had been spilt 58. The Rena was wrecked close to Tauranga, a major port and city with all the infrastructure required to contain the oil spill and treat oiled wildlife; no such facilities exist close to areas inhabited by tawaki. Foveaux Strait is an important passage for international shipping (http://bit.ly/foveaux-traffic), so that the region is more likely to experience a disaster than South Westland where there is little inshore boat traffic. In Fiordland, growing tourism numbers is leading to increased ship traffic; spills from large cruise vessels while in the fjords could severely impact local penguin populations. The risk of oil spills has been reduced after the recent termination of new offshore oil exploration leases. With the exception of one petroleum well some 50 km due west of Hokitika, there are currently no active oil exploration permits within the home range of breeding tawaki (http://bit.ly/petroleum-wells).
Fisheries interactions
Resource competition
South Westland: Minor
Fiordland: Minor
Foveaux Strait: Minor
Otley et al.34 discuss the potential for resource competition between commercial fisheries and tawaki. The authors conclude that due to the limited dietary information about tawaki, it is difficult to adequately assess competition between fisheries and penguins. That the penguins primarily target larval and juvenile stages of their prey species52,53 reduces the risk for direct competition with commercial fisheries. However, overfishing of prey stocks may reduce spawning biomass which could have an indirect effect on the penguins’ diet.
Incidental bycatch
South Westland: Minor
Fiordland: Major
Foveaux Strait: Major
In New Zealand, set net fisheries pose the single most significant cause of incidental mortality in penguins59. While the species has been assessed as having a low bycatch risk60, this assessment averages the risk across the entire tawaki breeding range. Furthermore, small vessel, inshore fisheries, those most likely to catch penguins, were not the focus of their analysis and these fisheries have very limited observer coverage. Thus, there is no robust estimate of tawaki bycatch. In 2016 two tawaki were killed in set nets in Dusky Sound in southern Fiordland59. Both birds drowned in nets targeting butterfish, a species that is primarily caught in kelp forests close to the coast, a habitat where tawaki forage. There is no set net ban in place around any of the islands in Foveaux Strait so that the risk of accidental bycatch of tawaki breeding in this region is real.
Marine predators
Otley et al.34 list inter-specific competition and predation by marine mammals as a threat. They note that recovering populations of Antarctic fur seals (Arctocephalus gazella) have top-down effects on Macaroni penguin (Eudyptes chrysolophus) populations. New Zealand fur seal (Arctocephalus forsteri) numbers in South Westland may in fact be declining34, suggesting that the impact of fur seals locally is likely to be negligible. Population trends for fur seals in Fiordland and Stewart Island are not known but increases are likely. Whether natural predators should (or would) be managed is a moot question, especially since any intervention is limited by the protective status of these species in New Zealand.
Terrestrial predators
Stoats
South Westland: Medium
Fiordland: Minor
Foveaux Strait: not a threat
Stoats can have a significant effect on the reproductive outcome of tawaki. There is little evidence to suggest that adult penguins are vulnerable to stoat predation34. But the loss of most tawaki eggs or chicks at Jackson Head caused by stoats in August and September 2016 shows the potential severity mustelid invasions39,40. The predatory pressure from stoats is highly variable – zero some seasons but almost total breeding failure in other years40 – and depends both on the availability of alternative prey (e.g. rats) as well as the abundance of stoats61. Mustelids are absent from those Foveaux Strait Islands where Tawaki breed and from some islands in South Westland (Open Bay Islands) and some sites in Fiordland.
Dogs
South Westland: Minor
Fiordland: not a threat
Foveaux Strait: not a threat
Uncontrolled dogs are still common in many of the West Coast’s remote communities and occasional kills of tawaki by stray dogs occurs. While this is an unnecessary and avoidable threat, it is unlikely that dog attacks will reach a significance that may have population-wide consequences.
Weka
South Westland: Minor
Fiordland: not a threat
Foveaux Strait: Minor
Weka can only be considered a minor threat where the birds have been introduced by humans, i.e. Open Bay Island and Solander Island. Elsewhere weka - itself classified a threatened species (nationally vulnerable13) - is a natural predator that has co-existed with tawaki since millennia62.
Disease
In the 1970s, an endemic blood parasite Leucocytozoon tawaki was isolated from tawaki with sandflies (Austrosimulium sp.) identified as the primary vector63,64. However, while Leucocytozoon appear to cause sporadic chick mortality in yellow-eyed penguins 65, there is no other published evidence of ill-effects of these parasites in other penguin populations66,67.
Avipox
South Westland: Minor
Fiordland: unknown
Foveaux Strait: unknown
Avipox is the only disease that has been observed to cause mortality in tawaki. So far, the virus has been reported only once in 2017 at a single site in South Westland and only affected a small number of chicks. In Magellanic penguins, fleas have been implicated as the primary vector for the virus68; it seems plausible that in tawaki sandflies (Austrosimulium sp.) may play this role.
Climate change
Ocean warming
South Westland: unknown
Fiordland: Minor
Foveaux Strait: unknown
Increasing sea surface temperatures have been implicated as a driving factor for population declines in crested penguin populations in New Zealand and elsewhere6,44,69,70. The population of Snares penguins has been stable throughout the last century despite significant variations in ocean temperatures71. With offshore foraging tawaki using similar subtropical water masses, it may be that tawaki are also is less affected by warming oceans than sub-Antarctic breeding crested penguins. However, the true impacts of warming oceans can only be assessed with further studies of their marine ecology and more reliable information about tawaki demography.
Weather extremes (E. Niño/La Niña)
South Westland: Major
Fiordland: inner fjords - not a threat, open coast - unkown
Foveaux Strait: unknown
The strong El Niño in 2015 had a significant impact on the breeding success of offshore foraging tawaki at Jackson Head38. Since the reversal of wind patterns and offshore dispersal of nutrients are believed to be responsible for the longer foraging ranges and lower foraging success in 2015 (see https://youtu.be/1c4B9sdL8Y0?&t=12m22s, it must be assumed that similar effects were apparent along the entire open coast of South Westland and Fiordland. Although the 2017 breeding season coincided with a strong La Niña, the substantially higher than normal ocean temperatures were not reflected in the penguins’ reproductive success. While still substantially lower when compared to Fiordland and Codfish Island/Whenua Hou penguins, many pairs at Jackson Head raised chicks to fledging. Nevertheless, foraging ranges were longer than during ‘normal’ years (Mattern, unpubl. data) indicating La Niña did affect the birds, albeit not as severely as El Niño. In Fiordland, the effect of El Niño appeared to be diametrically different with high breeding success and short foraging ranges that were little different to those in 2014 and 2016, indicating that the weather phenomena had no impact on Milford penguins. While no information about foraging behaviour or breeding success is available for Codfish Island/Whenua Hou during the 2015 El Niño, tawaki there performed trips during the 2017 La Niña that were comparable to 2016, when the environmental conditions were average. So, it seems that the penguins may have adequate coping mechanisms reducing the potential impact of weather extremes. However, as with ocean warming, these conclusions are based on few observations and further information is required.
Human interactions
Unregulated visitation
South Westland: Medium
Fiordland: Minor
Foveaux Strait: Minor
Tawaki breed in areas that are mostly inaccessible to visitors. Nevertheless, a few tawaki breeding sites, in particular Monro Beach and to a lesser extent Jackson Head, receive frequent, largely unregulated visits from tourists, where people on the beach may deter tawaki from landing and, hence, disrupt provisioning of their chicks. This in turn, could reduce reproductive success and eventually a decline of local penguin numbers. A recent problem is disturbance of moulting tawaki by people wanting to take photographs or ‘selfies’ with penguins. These incidents usually only affect a few individuals and have little or no population-wide effect. The Wilderness Lodge at Lake Moeraki hold the only concession to take tourists to a nearby beach. Their visits are well regulated and pose no threat to the penguins, however, other less well-behaved people sometimes visit this colony (K-J. Wilson, pers. obs.).
Boat traffic
South Westland: Minor
Fiordland: Medium
Foveaux Strait: Minor
While there are currently no obvious deleterious effects of boat traffic in Milford Sound/Piopiotahi, projected visitor numbers suggest that a substantial increase in the number of boats cruising the fjord is likely. Many of the boats drive close to the cliff walls on either side of the fjord where the penguins forage. This may pose a risk from prop strikes for diving penguins72. Although the effects of increasing underwater noise are not understood, it could have detrimental effects on both penguins and the distribution of their prey. There is little information about boat traffic in other fjords. Observations of fishing vessels in the southern fjords in the 1970s showed that many skippers were not considerate of wildlife (K.-J. Wilson, pers. obs.). If this still applies to the newer generation of fishermen and whether it poses a significant threat to local tawaki populations is unknown.
A proposed water pipeline and marine loading facility at Jackson Head could pose a threat to the local penguin population; particularly by stirring up of sediments that then drift towards Jackson Head reducing underwater visibility and compromise the penguins’ ability to find food or make landfall.
Research Priorities
While recent research has advanced out knowledge about tawaki, the species remains one of the least known of the world’s penguins. The demographic work in particular has raised more questions than answers. It is imperative to develop enhanced monitoring methods to get a better understanding of population trends.
1. Population monitoring
| R.1.H1 |
Investigate viability of automated monitoring solutionsUsing nest counts to acquire reliable data on demographic parameters has proven to be very difficult. Otley et al. (2017) suggest that the use of automated monitoring solutions (e.g. transponder gateways) would be impracticable for the use with tawaki due to the birds unpredictable landing sites. However, most breeding sites have access paths that are used by a large portion of the local penguin population. Therefore, automated identification systems using transponder tagged penguins may be a more reliable – and less resource demanding – method to assess population trends. Establish transponder marked tawaki subpopulations that can be monitored permanently via an automated transponder reading setup. Viable sites could be Gorge River, Milford Sound, Doubtful Sound and Codfish Island/Whenua Hou; all sites have a more or less permanent human presence which would allow for maintenance of the system. |
| R.1.H2 |
Establish marked populations at sites representative of the species diverse habitat (i.e. West coast, inner & outer Fiordland, Foveaux Strait)Using subcutaneous transponders, penguins can be marked with no impact on their hydrodynamics and the necessity for frequent band maintenance. At Jackson Head, Harrison Cove in Milford Sound/Piopiotahi, and on Codfish Island/Whenua Hou many penguins are already marked. In conjunction with automated monitoring gates, these marked populations would probably be a superior way to assess population trends than nest counts. However, each year, fledglings need to be marked. Hence, accessible sites should be chosen. Establish transponder marked tawaki subpopulations that can be monitored permanently via an automated transponder reading setup. Viable sites could be Gorge River, Milford Sound, Doubtful Sound and Codfish Island/Whenua Hou; all sites have a more or less permanent human presence which would allow for maintenance of the system. |
| R.1.H3 |
Conduct ground surveys in Fiordland and Stewart Island to establish occupancy densitiesWhile ground surveys are not suitable to reliably determine true numbers of breeding pairs, it is essential to establish where tawaki colonies are located. Of importance are the inside of the fjords and Stewart Island coast, where there is little information on penguin distribution. Penguin colonies should be mapped in detail to assess future changes. Survey coastal regions of the major fjords and Stewart Island during the chick rearing period where penguin vocalisations can be used to detect breeding colonies. Ground surveys should be conducted by experienced observers. |
| R.1.H4 |
Monitor breeding successDetermining the actual breeding success in tawaki is difficult since chicks tend to move around the colony during the crèching stage. As a result, the likelihood of finding older chicks at their nest sites is reduced towards the end of the breeding season. Develop better ways to monitor breeding success. Marking chicks early in crèching when they tend to return to the nest at night with transponders may be a viable option if used in combination with automated monitoring systems (see above) counting the number that eventually depart seawards. Another option could be to use time-lapse cameras that record penguin traffic on the beach during the fledging period and provide a measure of number of chicks that eventually leave the colony. |
| R.1.H5 |
Examine regional variation in threats to survival (e.g. set net bycatch, predator presence) and breeding failure. |
| R.1.M6 |
Investigate the potential for range expansion of the speciesConduct regular surveys of areas suitable for, but not currently recorded as breeding habitat (e.g. Catlins, Otago Peninsula). |
| R.1.M7 |
Examine the prevalence and distribution of disease pathogens and vectors throughout the species’ breeding range.Particular emphasis should be on avian pox which caused the death of a chick on the West Coast and avian malaria which affects yellow-eyed penguins. |
2. Marine Ecology
| R.2.H1 |
Examine the effects of environmental perturbations on the species across its diverse range.The substantially different effects of El Niño and La Niña on the foraging behaviour and success of penguins from coastal colonies compared to those foraging within a fjord have already been demonstrated. However, the environmental mechanisms underlying these differences are poorly understood. Could it be that fjords provide a safe haven for the species that buffers tawaki from major environmental perturbations? Does this also apply to tawaki from the Foveaux Strait region? Or are there regional differences between the coastal colonies along South Westland and the outer coast of Fiordland? |
| R.2.H2 |
Study differences in foraging behaviour of penguins breeding deep inside fjords with conspecifics from the outer coastlines of Fiordland to assess habitat characteristics, influence on species distribution, and buffer capabilities in the face of environmental change.Essentially an extension of R.2.H1. Do tawaki breeding at the interface of fjord and open sea environments benefit from the ability to switch between foraging in one or other environment depending on environmental conditions? |
| R.2.H3 |
Investigate fisheries impact.The majority of the species breeding range is exempt from the set-net ban that covers most of the South Island’s coastline. With their near-shore foraging strategies tawaki are prone to accidental entanglement in set net operations targeting butterfish. Trial and establish video monitoring of near shore set netters to assess the frequency of tawaki-fisheries interactions and, if necessary, develop mitigation measures. |
| R.2.H4 |
Monitor pre-moult and non-breeding dispersal.The pre-moult period is probably the most critical period in any migratory penguin species, especially if the penguins have just completed a resource demanding breeding phase. Hence, foraging success during the pre-moult dispersal is likely to be crucial for annual adult survival. With the apparent enormous distances travelled during this period, the consequences of environmental perturbations at the penguins’ travel destinations may have significant effects on survival. A multi-year, multi-site examination of pre-moult dispersal would provide insights into ways habitat variability affects penguin behaviour and survival. Track tawaki annual pre-moult dispersal using geolocator devices (GLS) that can be deployed for long periods (up to 5 years, minimal maintenance) at key sites (i.e. South Westland, Fiordland, Foveaux Strait). Examine whether this could also be achieved with GPS dive loggers (i.e. similar to Whitehead et al.73). We do not know where tawaki go between moult and breeding. Tracking tawaki over their winter dispersal will show whether the penguins are exposed to different threats from those suffered in their breeding range and provide further information about environmental variables dictating their movement patterns and survival. Track tawaki through their winter dispersal with GLS loggers and satellite transmitters every 3-5 years. |
| R.2.H5 |
Establish a marine ecology monitoring programme during the breeding season.With the limited information about tawaki’s marine ecology, there is still a dearth of data to examine how environmental change may affect the species in its different foraging habitats. With advances in tracking technology increasing what aspects of penguins’ at-sea behaviour can be recorded, establishing a regular monitoring protocol for comparing changes in foraging behaviour between sites and over time is now viable. Similar programmes have been established for little penguins in Australia which have significantly advanced understanding of environmental impacts and subsequent conservation responses74,75. Deploy GPS dive loggers on 10-15 birds each year at three reference sites throughout the species range and assess annual variations in foraging parameters in response to environmental variables. |
| R.2.H6 |
Examine diet composition.We know very little about tawaki diet. What the penguins eat inside of fjord ecosystems needs to be investigated. Further information is required on foods from South Westland and the Foveaux Region. While stomach flushing has been found not to adversely affect penguins76, less intrusive and less laborious methods maybe more suitable for tawaki. Develop a database (or add to GenBank) of genetic barcodes for all potential prey species (including invertebrates that might not be detected with more traditional approaches). Use DNA faecal samples to determine prey composition at a range of sites. Examine viability of animal-borne cameras to examine prey encounter rates, pursuit strategies and number of preys taken. Collect feather samples from every handled tawaki to establish a feather database for later analysis of diet composition trends using stable isotope analysis. |
| R.2.M7 |
Examine effects of Marine Reserves in Fiordland on foraging movements.In Milford Sound/Piopiotahi, tawaki forage predominantly along the northern fjord boundaries within the Marine Reserve. Do the penguins forage in these regions as a result of environmental benefits due to the Marine Reserve (e.g. no holding pots)? |
Acknowledgments
We are grateful to Tess Cole, Helen Otley, Gerry McSweeney and Dave Houston for valuable feedback and criticism of earlier drafts of this account. This review was funded by the T-Gear Charitable Trust and the Birds-New Zealand Research Fund - thanks so much for supporting our work. We are especially grateful to Peter Gaze for his support and interest throughout.
References
- BirdLife International. (2018) Eudyptes pachyrhynchus. The IUCN Red List of Threatened Species 2018 e.T22697776A132601937 DOI: https://doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22697776A132601937.en
- Warham, J. (1974) The Fiordland Crested Penguin Eudyptes pachyrhynchus. Ibis 116: 1–27 DOI: https://doi.org/10.1111/j.1474-919X.1974.tb00220.x
- Marchant, S & Higgins, PJ. (1990) Eudyptes pachyrhynchus, Fiordland Penguin. in Handbook of Australian, New Zealand and Antarctic Birds. Volume 1: Ratites to Ducks. 195–205 (Oxford University Press).
- McLellan, R. (2009) Tawaki Monitoring Review and Analysis. West Coast Conservancy, Department of Conservation. Hokitika, New Zealand. Available at: https://ref.coastalrestorationtrust.org.nz/site/assets/files/8654/wildlands_tawaki_report_-_kaes_comments.pdf.
- Mattern, T. (2013) Fiordland penguin (Eudyptes pachyrhynchus). in Penguins: Natural History and Conservation (eds. García‐Borboroglu, P. & Boersma, P. D.) 152–167 (University of Washington Press).
- Taylor, GA. (2000) Action Plan for Seabird Conservation in New Zealand. Part A: Threatened Seabirds. Department of Conservation, New Zealand. Wellington.
- McLean, IG, Kayes, SD, Murie, JO, Davis, LS & Lambert, DM. (2000) Genetic monogamy mirrors social monogamy in the Fiordland crested penguin. New Zealand Journal of Zoology 27: 311–316
- Studholme, BJS. (1994) Parent-offspring recognition in the Fiordland Crested penguin (Eudyptes pachyrhynchus). New Zealand Natural Sciences 21: 27–36
- Grau, CR. (1982) Egg formation in Fiordland Crested Penguins (Eudyptes pachyrhynchus). The Condor 84: 172-177. Available at: https://sora.unm.edu/sites/default/files/journals/condor/v084n02/p0172-p0177.pdf
- McQueen, SM, Davis, LS & Young, G. (1998) The reproductive endocrinology of Fiordland Crested Penguins Eudyptes pachyrhynchus. Emu 98: 127–131 DOI: https://doi.org/10.1071/MU98013
- St Clair, CC. (1992) Incubation behavior, brood patch formation and obligate brood reduction in Fiordland crested penguins. Behavioral Ecology and Sociobiology 31: 409–416
- McLean, IG. (1990) Chick expulsion by a Fiordland Crested Penguin. Notornis 37: 181–182 Available at: http://notornis.osnz.org.nz/chick-expulsion-fiordland-crested-penguin.
- Robertson, HA et al. (2017) Conservation status of New Zealand birds, 2016. New Zealand Threat Classification Series 19: 26 p. Available at: http://www.doc.govt.nz/upload/documents/science-and-technical/nztcs4entire.pdf.
- Kinsky, FC. (1970) Annotated checklist of the birds of New Zealand including the birds of the Ross Dependency. (Reed).
- Stonehouse, B. (1971) The Snares Islands Penguin Eudyptes robustus. Ibis 113: 1–7
- Falla, RA, Warham, J & Fleming, CA. (1974) Comments on the proposed preservation of Eudyptes sclateri Buller, 1888 and Eudyptes robustus (Oliver, 1953). Bulleting of Zoological Nomenclature 30: 136
- Warham, J. (1974) The breeding biology and behaviour of the snares crested penguin. Journal of the Royal Society of New Zealand 4: 63–108 DOI: https://doi.org/10.1080/03036758.1974.10419407
- Baker, AJ, Pereira, SL, Haddrath, OP & Edge, K-A. (2006) Multiple gene evidence for expansion of extant penguins out of Antarctica due to global cooling. Proceedings of the Royal Society B: Biological Sciences 273: 11–7 DOI: https://doi.org/10.1098/rspb.2005.3260
- Ksepka, DT, Bertelli, S & Giannini, NP. (2006) The phylogeny of the living and fossil Sphenisciformes (penguins). Cladistics 22: 412–441 DOI: https://doi.org/10.1111/j.1096-0031.2006.00116.x
- Davis, LS & Renner, M. (2003) Penguins. (T&AD Poyser).
- Cole, TL et al. (2019) Ancient DNA of crested penguins: Testing for temporal genetic shifts in the world’s most diverse penguin clade. Molecular Phylogenetics and Evolution 131: 72–79 DOI: https://doi.org/10.1016/j.ympev.2018.10.025
- Christidis, L & Boles, W. (2008) Systematic and taxonomy of Australian birds. CSIRO Publishing Available at: http://www.publish.csiro.au/book/5702/%0Ahttps://academic.oup.com/sysbio/article-lookup/doi/10.1093/sysbio/syp071.
- Tobias, JA et al. (2010) Quantitative criteria for species delimitation. Ibis 152: 724–746 DOI: https://doi.org/10.1111/j.1474-919X.2010.01051.x
- Holdaway, RN, Worthy, TH & Tennyson, AJD. (2001) A working list of breeding bird species of the New Zealand region at first human contact. New Zealand Journal of Zoology 28: 119–187 DOI: https://doi.org/10.1080/03014223.2001.9518262
- Worthy, TH. (1997) The identification of fossil Eudyptes and Megadyptes bones at Marfells Beach, Marlborough, South Island. New Zealand Natural Sciences 23: 71–85
- Mattern, T et al. (2018) Marathon penguins – Reasons and consequences of long-range dispersal in Fiordland penguins / Tawaki during the pre-moult period. PLOS ONE 13: e0198688 DOI: https://doi.org/10.1371/journal.pone.0198688
- McLean, IG & Russ, RB. (1991) The Fiordland Crested Penguin survey, stage I: Doubtful to Milford sounds. Notornis 38: 183–190
- Russ, RB, McLean, IG & Studholm, BJS. (1992) The Fiordland Crested Penguin survey, stage II: Dusky and Breaksea sounds. Notornis 39: 113–118
- McLean, IG, Studholm, BJS & Russ, RB. (1993) The Fiordland Crested Penguin survey, stage III: Breaksea Island, Chalky and Preservation inlets. Notornis 40: 85–94
- Studholm, BJS, Russ, RB & McLean, IG. (1994) The Fiordland Crested Penguin survey: stage IV, Stewart and offshore islands and Solander Island. Notornis 42: 133–143
- McLean, IG et al. (1997) The Fiordland Crested Penguin (Eudyptes pachyrhynchus) survey, stage V: mainland coastline, Bruce Bay to Yates Point. Notornis 44: 37–47
- Mattern, T & Long, R. (2017) Survey and population size estimate of Fiordland penguin / tawaki (Eudyptes pachyrhynchus) in Milford Sound / Piopiotahi, New Zealand. Notornis 64: 97–101
- Long, R. (2017) A survey of Fiordland crested penguins / tawaki (Eudyptes pachyrhynchus) from Cascade River to Martins Bay, South Westland, New Zealand, 2014. Notornis 64: 206–210
- Otley, H et al. (2018) Assessing the population trend and threats to New Zealand’s Fiordland crested penguin using counting and demographic modelling approaches. New Zealand Journal of Ecology 42: DOI: https://doi.org/10.20417/nzjecol.42.15
- Young, M, Pullar, C & McKinlay, B. (2015) Breeding attempts by Fiordland crested penguins/tawaki (Eudyptes pachyrhynchus) on the Otago Coast. Notornis 62: 102–104
- St. Clair, CC. (1998) Eudyptes pachyrhynchus. in Report from a workshop held 8-9 September 1996, Cape Town, South Africa (eds. Ellis, S., Croxall, J. P. & Cooper, J.) 69–72 (IUCN/SSC Conservation Breeding Specialist Group).
- Ellenberg, U et al. (2015) Assessing the impact of nest searches on breeding birds-a case study on Fiordland crested penguins (Eudyptes pachyrhynchus). New Zealand Journal of Ecology 39: 231
- Mattern, T & Ellenberg, U. (2016) The Tawaki Project: Field Report 2015 - year 2, 13 September-14 October 2015. NZ Penguin Initiative. Dunedin, NZ. DOI: https://doi.org/10.13140/RG.2.1.3068.1847
- Mattern, T & Ellenberg, U. (2017) The Tawaki Project - Field Report 2016 - Year 3, 16 September - 13 November 2016. NZ Penguin Initiative. Dunedin, New Zealand. DOI: https://doi.org/10.13140/RG.2.2.31050.77760
- Wilson, K-J & Long, R. (2018) Tawaki / Fiordland Crested Penguin Conservation Management; Predator control –four year final report, 2014-2017. Hokitika, New Zealand.
- Long, RG, Long, RB, Stewart, CA & Otley, H. (2009) Surveys for Fiordland crested penguin from Sandrock Bluff to Longridge Point, South Westland, August and September 2009 (DOCDM-562482). Wellington, New Zealand.
- Long, RG, Long, RB & Stewart, CA. (2011) Penguin survey of Tawaki Fiordland crested penguin. A report to West Coast Penguin Trust and DOC Hokitika (DOCDM-898531). Wellington, N.Z.
- Otley, H, Tansell, J & Scofield, P. (2017) A comprehensive demographic assessment of the endangered Fiordland crested penguin Eudyptes pachyrhynchus. New Zealand Journal of Zoology 1–19 DOI: https://doi.org/10.1080/03014223.2017.1284135
- Morrison, KW, Battley, PF, Sagar, PM & Thompson, DR. (2015) Population dynamics of Eastern Rockhopper Penguins on Campbell Island in relation to sea surface temperature 1942–2012: current warming hiatus pauses a long-term decline. Polar Biology 38: 163–177
- Guinard, E, Weimerskirch, H & Jouventin, P. (1998) Population Changes and Demography of the Northern Rockhopper Penguin on Amsterdam and Saint Paul Islands. Colonial Waterbirds 21: 222–228 DOI: https://doi.org/10.2307/1521909
- Dehnhard, N, Poisbleau, M, Demongin, L, Ludynia, K & Quillfeldt, P. (2014) High juvenile annual survival probabilities in Southern Rockhopper Penguins Eudyptes chrysocome are independent of individual fledging traits. Ibis 156: 548–560 DOI: https://doi.org/10.1111/ibi.12167
- St Clair, CC. (1999) Fidelity to nest site and mate in fiordland crested penguins Eudyptes pachyrhynchus. Marine Ornithology 27: 40–41
- Warham, J. (1975) The crested penguins. in The biology of penguins (ed. Stonehouse, B.) 189–269 (Macmillan).
- Mattern, T & Ellenberg, U. (2018) The Tawaki Project: Field Report 2017 - year 4, 16 September-02 March 2015. NZ Penguin Initiative. Dunedin, NZ. DOI: https://doi.org/10.13140/RG.2.2.12459.95524/2
- St. Clair, CC & St. Clair, RC. (1992) Weka predation on eggs and chicks of Fiordland crested penguins. Notornis 39: 60–63
- Mattern, T & Ellenberg, U. (2015) The Tawaki Project: Field Report 2014 - Year 1, 26 August-17 November 2014. NZ Penguin Initiative. Dunedin, New Zealand. DOI: https://doi.org/10.13140/RG.2.1.4116.7602
- van Heezik, Y. (1990) Diets of yellow-eyed, Fiordland crested, and little blue penguins breeding sympatrically on Codfish Island, New Zealand. New Zealand Journal of Zoology 17: 543–548
- van Heezik, Y. (1989) Diet of Fiordland Crested penguin during the post-guard phase of chick growth. Notornis 36: 151–156
- Mattern, T & Ellenberg, U. (2018) Yellow-eyed penguin diet and indirect effects on prey composition – Collation of biological information (CSP16205-1, POP2016-05). (Department of Conservation, New Zealand). DOI: https://doi.org/10.13140/RG.2.2.23828.81284
- Watanuki, Y et al. (2006) Swim speeds and stroke patterns in wing-propelled divers: a comparison among alcids and a penguin. Journal of Experimental Biology 209: DOI: https://doi.org/10.1242/jeb.02128
- Biasi, GP, Langridge, RM, Berryman, KR, Clark, KJ & Cochran, UA. (2015) Maximum‐Likelihood Recurrence Parameters and Conditional Probability of a Ground‐Rupturing Earthquake on the Southern Alpine Fault, South Island, New Zealand. Bulletin of the Seismological Society of America 105: 94–106 DOI: https://doi.org/10.1785/0120130259
- Jones, H, Poot, M, Mullarney, J, de Lange, W & Bryan, K. (2016) Oil dispersal modelling: reanalysis of the Rena oil spill using open-source modelling tools. New Zealand Journal of Marine and Freshwater Research 50: 10–27 DOI: https://doi.org/10.1080/00288330.2015.1112819
- Schiel, D, Ross, P & Battershill, C. (2016) Environmental effects of the MV Rena shipwreck: cross-disciplinary investigations of oil and debris impacts on a coastal ecosystem. New Zealand Journal of Marine and Freshwater Research 50: 1–9 DOI: https://doi.org/10.1080/00288330.2015.1133665
- Crawford, R et al. (2017) Tangled and drowned: A global review of penguin bycatch in fisheries. Endangered Species Research 34: 2017 DOI: https://doi.org/10.3354/esr00869
- Richard, Y & Abraham, ER. (2015) Assessment of the risk of commercial fisheries to New Zealand seabirds, 2006–07 to 2012–13. Wellington, New Zealand. Available at: https://www.mpi.govt.nz/document-vault/10523.
- Tompkins, DM, Byrom, AE & Pech, RP. (2013) Predicted responses of invasive mammal communities to climate-related changes in mast frequency in forest ecosystems. Ecological Applications 23: 1075–1085 DOI: https://doi.org/10.1890/12-0915.1
- Beauchamp, AJ. (1989) Pan biogeography and rails of the genus Gallirallus. New Zealand Journal of Zoology 16: 763–772
- Fallis, AM, Bisset, SA & Allison, FR. (1976) Leucocytozoon tawaki n.sp. (Eucoccida: Leucocytozoidae) from the penguin Eudyptes pachyrhynchus, and preliminary observations on its development in Austrosimulium spp. (Diptera: Simuliidae). New Zealand Journal of Zoology 3: 11–16
- Allison, FR, Desser, SS & Whitte, LK. (1978) Further observations on the life cycle and vectors of the haemosporidian Leucocytozoon tawaki and its transmission to the Fiordland crested penguin. New Zealand Journal of Zoology 5: 371–374
- Argilla, LS. (2015) An investigation of the causes of mortality in yellow-eyed penguins (Megadyptes antipodes) across their range with specific emphasis on the role played by Leucocytozoon. (Massey University, Palmerston North, New Zealand).
- Jones, HI & Shellam, GR. (1999) Blood parasites in penguins, and their potential impact on conservation. Marine Ornithology 27: 183–184
- Vanstreels, RET, Braga, ÉM & Catão-Dias, JL. (2016) Blood parasites of penguins: A critical review. Parasitology 143: 931–956 DOI: https://doi.org/10.1017/S0031182016000251
- Kane, OJ et al. (2012) Avian Pox in Magellanic Penguins (Spheniscus magellanicus). Journal of Wildlife Diseases 48: 790–794 DOI: https://doi.org/10.7589/0090-3558-48.3.790
- Cunningham, DM & Moors, PJ. (1994) The Decline of Rockhopper Penguins Eudyptes chrysocome at Campbell Island, Southern Ocean and the Influence of Rising Sea Temperatures. Emu 94: 27–36
- Hilton, GM et al. (2006) A stable isotopic investigation into the causes of decline in a sub-Antarctic predator, the rockhopper penguin Eudyptes chrysocome. Global Change Biology 12: 611–625
- Mattern, T, Houston, DM, Lalas, C, Setiawan, AN & Davis, LS. (2009) Diet composition, continuity in prey availability and marine habitat – keystones to population stability in the Snares Penguin (Eudyptes robustus). Emu 109: 204–213 DOI: https://doi.org/10.1071/MU0800
- Cannell, BL et al. (2016) Anthropogenic trauma is the most prevalent cause of mortality in Little Penguins, Eudyptula minor, in Perth, Western Australia. Emu 116: 52–61 DOI: https://doi.org/http://dx.doi.org/10.1071/MU15039
- Whitehead, TO, Kato, A, Ropert-Coudert, Y & Ryan, PG. (2016) Habitat use and diving behaviour of macaroni Eudyptes chrysolophus and eastern rockhopper E. chrysocome filholi penguins during the critical pre-moult period. Marine Biology 163: 1–20 DOI: https://doi.org/10.1007/s00227-015-2794-6
- Saraux, C, Chiaradia, A, Salton, M, Dann, P & Viblanc, VA. (2016) Negative effects of wind speed on individual foraging performance and breeding success in little penguins. Ecological Monographs 86: 61–77 DOI: https://doi.org/10.1890/14-2124.1
- Pelletier, L, Chiaradia, A, Kato, A & Ropert-Coudert, Y. (2014) Fine-scale spatial age segregation in the limited foraging area of an inshore seabird species, the little penguin. Oecologia 176: 399–408 DOI: https://doi.org/10.1007/s00442-014-3018-3
- Goldsworthy, B, Young, MJ, Seddon, PJ & van Heezik, Y. (2016) Stomach flushing does not affect apparent adult survival, chick hatching, or fledging success in yellow-eyed penguins (Megadyptes antipodes). Biological Conservation 196: 115–123 DOI: https://doi.org/10.1016/j.biocon.2016.02.009